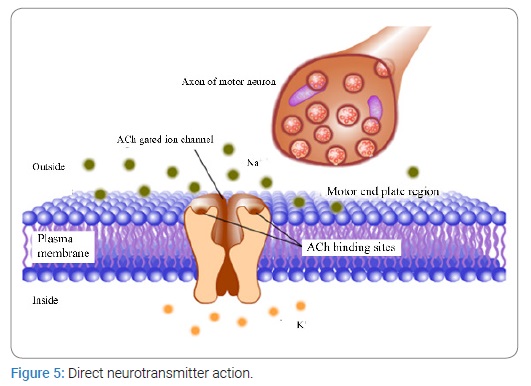
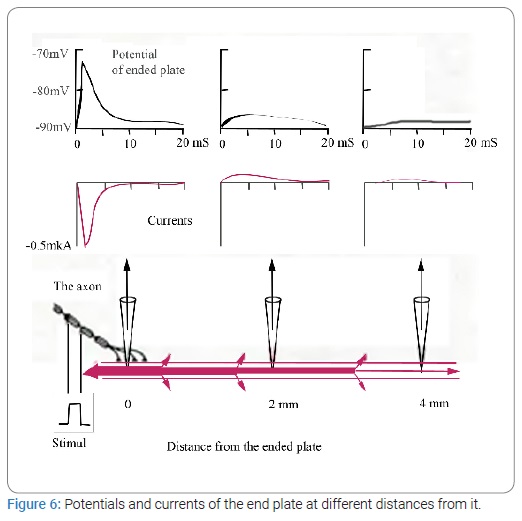
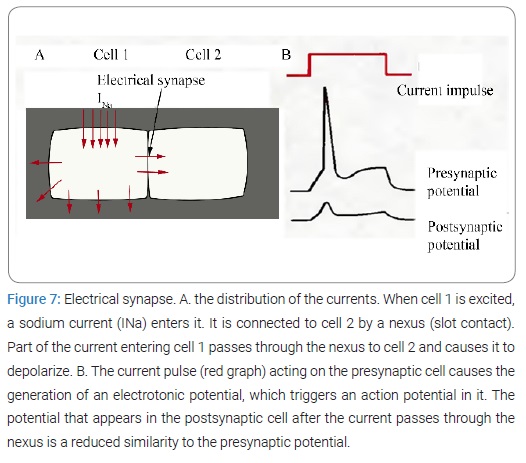
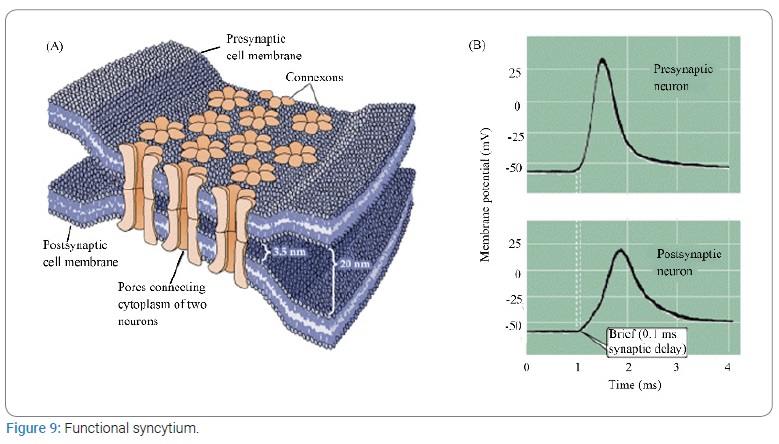
An electric current, Electrical Stimulation of the Brain (ESB), and its effect on behavioral process
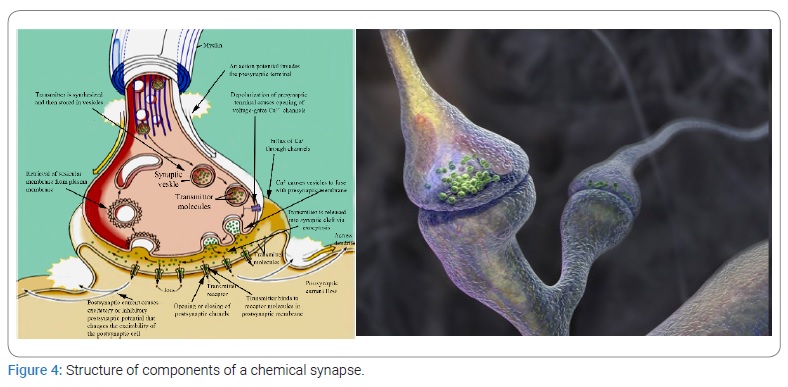
There is a huge gap between our knowledge of electrical phenomena at the cellular level and our understanding of the mechanisms underlying responses of the whole body. How can activities such as walking, problem solving, or thinking is explained in terms of cell membrane polarization and repolarization? Of course, behavioral reactions are impossible without corresponding spike potentials and ion exchange, but they are also impossible without the consumption of sugars and oxygen; therefore, we must distinguish the main nonspecific processes of cell activity—cellular metabolism—from processes specifically related to behavior. Electrical activity of neurons seems to play a dual role: on the one hand, it is a manifestation of cell metabolism, and on the other, it is a way to transmit encoded information. The ability to transmit encoded information is the most important and least understood property of a neuron, i.e., a functional unit of neural communication.
Our knowledge on the physical and chemical foundations of biological activity, which have been addressed in many works [5–7], allows us to formulate the following postulates: 1) no behavioral act, including its mental components, is possible without the emergence of negative potentials that are accompanied by electrical and chemical shifts at the cellular level; 2) the membrane depolarization artificially caused by an electric current or a chemical substance can be followed by a behavioral reaction detectable by an observer; 3) even though behavior is very complex and many of its parameters are insufficiently studied, they are governed by the laws of physical and chemical processes that can be studied experimentally.
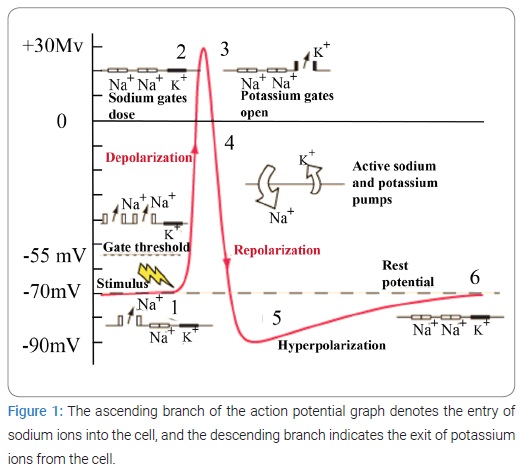
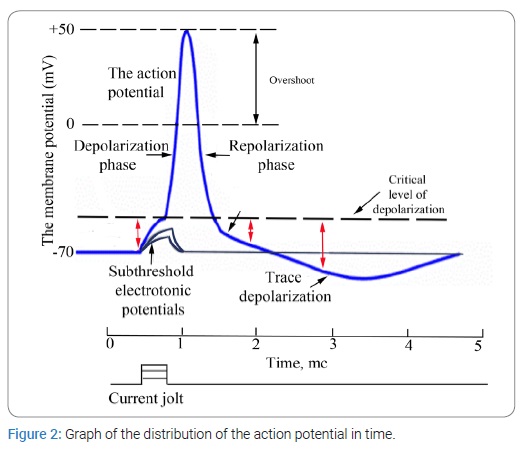
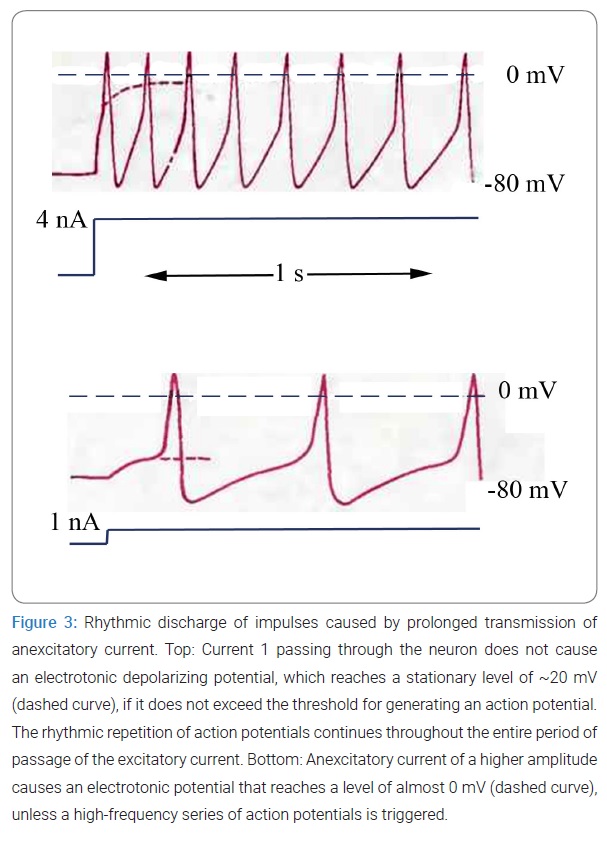
The apparent simplicity and constancy of muscle contraction can lead to erroneous conclusions because in fact contractility is an incredibly complex phenomenon involving. i) depolarization of the resting membrane; ii) changes in its permeability; iii) extremely rapid exchange of potassium, sodium, and other ions; iv) the emergence of electric fields; v) changes in the orientation of protein molecules in the muscle fiber accompanied by shortening of polypeptide chains; vi) decomposition and synthesis of adenosine triphosphate; vii) exchange of phosphate residues; viii) decomposition of hexose phosphate into lactic acid; and ix) many other biochemical reactions that proceed in an orderly fashion in the muscle fiber according to a genetically predetermined plan, regardless of the nature of the agent that caused them. The mechanisms of contraction and relaxation of the muscle fiber are embedded in the structure of its cells. An electric current, like the nervous system itself, only triggers all these processes. This principle is fundamental for understanding the control of biological functions by electricity.
In brain research, as in the study of muscle, it is best to use an electric current because it does not damage cells and allows an investigator to repeatedly analyze the same biological process. With the help of an electric current, we can activate the functional mechanisms inherent in cell structure and find out their possible involvement in spontaneous behavior. Using ESB, an investigator can control a variety of functions (cell movement or secretion by glands) or specific mental processes, depending on the purpose of the study. After the mastery of the technique of electrode implantation into the brain of animals, great progress has been made in the research on the impact of an electrical signal on animal behavior. In some chimpanzees, so many electrodes have been inserted into the brain that the total number of their contacts with the brain reached 100, and no neurophysiological or behavioral disorders were observed in this experiment; some monkeys managed to insert electrodes into such vitally important and finely organized structures as the respiratory center of the medulla oblongata. Furthermore, these experimental data were applied to a person. Experience has confirmed the safety of long-term implantation of electrodes into the human brain, and this method has long been used in specialized clinics in many countries around the world for the treatment of thousands of patients with epilepsy, involuntary movements, phantom pain, pathological feelings of anxiety, and other diseases of the central nervous system. Usually, several thin electrodes (a total of 20 contacts to 40 contacts) are implanted into surface or deep structures of the brain. In some patients, the electrodes have stayed in the brain for many years, without causing any adverse effects. Implantation of electrodes for a long period makes it possible to make a thorough time-limited diagnosis as well as to implement multiple electrical stimuli or strictly regulated electrocoagulation, depending on the nature of a disease. In addition, we can learn important information on the interdependence of mental and neurophysiological processes, thereby obtaining specific knowledge about the cerebral mechanisms of human behavior.
Experiments conducted by Jose Delgado [8], with Paddy the chimpanzee made it possible to investigate complex characteristics of behavior in the primate order. Paddy led a normal life in a cage, while on her head, a reinforced stimoceiver telemetrically transmitted electrical activity of her right and left amygdala nuclei to an adjacent room, where these signals were received, recorded, and automatically analyzed on a computer connected to this system. This machine was trained to recognize certain types of waves-so-called spindle flashes-that normally occur in both amygdala nuclei several times a minute and last ~1 s. The computer was also trained to turn on the transmitter, and as soon as the spindles appeared, radio signals were sent to Paddy’s brain, which stimulated a certain region of the reticular formation; the excitement at this site suppressed the spindle flashes. In this way, the emergence of a certain type of EEG pattern in one structure of the brain caused electrical stimulation of another structure, and the entire process of decoding information and sending signals was carried out by the computer. The results of the experiments revealed that 2 hours after feedback was established in the brain–computer–brain system, spindle flashes (reflecting the activity of the amygdala nuclei) occurred half as often, and after daily 2 hours sessions of electric stimulation for 6 days, this type of activity decreased to 1% of the normal frequency of its occurrence; at the same time, the animal became calmer and less alert and active when behavioral tests were conducted, although it continued without errors to perform tasks involving visual and olfactory analyzers. After that, the computer was shut down, and 2 weeks later, Paddy’s EEG and behavior returned to normal.
It should be noted that the scientific literature now contains quite a lot of data indicating amazing effects of ESB. For example, a researcher can stop the heart for a few moments, slow down or speed up its rhythm by appropriate stimulation of certain cortical and sub-cortical structures, and thus demonstrate that the brain controls the heart, and not the other way around. With the help of ESB, the researcher can change the frequency and amplitude of respiratory movements, peristalsis, and the secretory function of the stomach, arbitrarily adjust the diameter of the pupils from maximal constriction to maximal dilation, like the aperture of a camera, by simply changing the magnitude of electrical stimulation of the hypothalamic parts in the brain [9]. By means of ESB, an investigator can influence most of autonomic functions as well as sensory perception, motor activity, and mental functions. Instead of analyzing each experiment in detail, we chose a few typical examples to illustrate the main features of electrical control of the brain and the resulting behavioral changes.
Electrical stimulation of the motor cortex of the right hemisphere in a cat can cause the left hind leg to bend, and the amplitude of movement will be proportional to the applied current. For instance, in one of the experiments, when the animal’s initial position was on all four legs, a 1.2 mA current caused slight flexion of the paw, which was thus slightly lifted off the floor. When stimulated by a current of 1.5 mA, the paw rose 4 cm, and when the current was 1.8 mA, the flexion was complete. This movement developed gradually, reached its maximum within 2 seconds, and continued until the electric stimulation stopped. This movement could be repeated any number of times and was accompanied by adaptation of the entire body posture of the animal, including lowering of the head, raising of the pelvis, and slight shifting of the center of gravity to the left, in order to maintain balance while standing on three legs. The electrical stimulation did not cause any emotional disturbances: the cat remained as active and friendly as ever, rubbing its head against the experimenter, looking for human affection and purring.
Another one of Delgado’s elegant studies was conducted to investigate the effect of ESB on aggressive animal behavior. This study was carried out in a monkey colony with changes in its composition to gradually raise the social position of one of the animals, a female named Lina, who in the first group, consisting of four monkeys, occupied the lowest position, in the second group was the third, and in the third group, the second. The social position of the animal in a group was evaluated during long control periods using the following criteria: frequency of spontaneous approaches and sexual intercourse, the order of food intake, and a location occupied in a cage. In each of the three groups, for 2 days in a row, radio stimulation was applied to the posterolateral nucleus in Lina’s visual tubercle for 5 seconds every minute; the stimulation session was performed in the morning and lasted for 1 hour. In all cases, the brain stimulation caused Lina to run around the cage, climb the ceiling, lick her lips, emit screams, and, depending on her position in the group, attack other animals. In group 1, where Lina had a subordinate position, it tried to attack another monkey only once, whereas it was threatened and attacked 24 times. In group 2, she became more aggressive (24 attacks), and was attacked only 3 times, and in group 3, Lina attacked other monkeys 79 times and was not threatened once. In no group was there any change in the frequency of approaches between animals before or after the experiments; this shows that Lina’s aggressiveness was caused by an electric shock to the brain. These studies by Jose Delgado perfectly illustrate the crucial role of electrical stimuli in all physiological processes, both simple and highly organized ones.
A great deal of research on the effects of electrical stimuli on various functions of the human body has been conducted in Lebedev’s laboratory. He has shown that the use of Transcranial Electrical Stimulation (TES) by means of a thermal power unit affects many parameters of fundamental systems of the mammalian body. Let us take a look at the main findings.
An analgesic effect: To obtain an analgesic effect, optimal parameters of the electrical stimulation were selected at the beginning of the experiments. The analgesic regimen was found in wide-ranging screening experiments on various animal species with the registration of the intensity of different types of reactions to acute pain or their timing in response to amniotic pain stimuli (pain tolerance); pain stimuli of different physical nature were applied.
At the first stage, the efficiency of electrical parameters was assessed, and among them, output was stabilized by voltage. Rectangular pulses were employed with a frequency between 50 Hz and 3.5 kHz and duration between 0.1 ms and 1.0 ms in the presence or absence of an additional constant component. The electrodes were placed on the forehead and behind the ears. A very weak analgesic effect was observed in rabbits receiving TES alone with the following parameters: frequency within 70 Hz to 80 Hz, pulse duration of 1 ms in the presence of an additional constant component.
In further experiments, it was found that the most pronounced analgesic effect takes place in rabbits at the stimulation frequency of 77 Hz, pulse duration 3.5 ms to 4.0 ms, at the 1:2 ratio of the pulse to the additional constant current component. When the rectangular pulse was replaced with a packet of pulses of the same duration with a filling frequency of 10 kHz (meander), the analgesic effect was preserved, and the local reflecting effect of the current somewhat diminished.
Thus, it turned out that, in contrast to the accepted concepts, the analgesic effect is caused by the thermal power unit within a very narrow range of parameters. It should be added that the analgesic effect was detected only when the applied current had a sagittal direction. No analgesic effect was observed with bilateral application of the electrodes.
It was also demonstrated that the optimal analgesic frequency is species-specific, although the general pattern of the “frequency–intensity of analgesia” relation is approximately the same among all studied animal species. It resembles a quasi-resonant curve.
Readers can see that when the frequency deviated by 2 Hz to 4 Hz from the optimal one, the analgesic effect almost disappeared. In clinical studies, it was also noted that the optimal analgesic frequency for a person is close to 77 Hz, and when it is shifted within 10%, the analgesic effect decreases sharply. The “pulse duration–analgesia intensity” relation at the optimal frequency is also described by a quasi-resonant curve, but its steepness is much lower, and the optimal point has no species specificity.
In experiments involving TPP generators with optimal output pulse characteristics in terms of frequency, duration, and voltage-stabilized output, it was shown that the presence of a substantial additional constant component is of great importance for the implementation of the analgesic effect. Nonetheless, this approach significantly increased the effective value of the current. In comparative studies, it has been found that when a TPP generator with current-stabilized output is utilized, the corresponding analgesic effect may be achieved at the same current value of the pulse current but without an additional constant component, i.e., when the current value was reduced threefold.
The authors of that study advanced hypotheses about the possible mechanisms of analgesic and conjugated effects of TES during this type of stimulation. In the late 1960s, in studies on the mechanisms of electronarcosis, it was shown that direct electrical stimulation of some medially located structures of the brain stem (hypothalamic nuclei, periaqueductal gray matter of the midbrain, bridge seam nuclei, and the medulla oblongata) can induce severe analgesia in animals [10]. The same was observed when these structures were stimulated through implanted electrodes in humans. Subsequently, this analgesia was termed stimulation analgesia, and the system of structures that implemented it was called the Antinociceptive System (ANS).
Extensive studies on the ANS have revealed that this system is involved not only in the regulation of pain sensitivity and conduction of pain impulses in the CNS but also in homeostatic regulation (normalization) of a number of physiological functions when disturbed. A considerable part of the ANS uses endogenous morphine like substances (endorphins), serotonin, and some other compounds (for example, cholinergic) such as neurotransmitters and neuromodulators [11].
It has been suggested that TES in the advanced mode is a noninvasive analog of stimulation analgesia. In this regard, the following questions were raised when the TPP mechanisms of action were investigated:
- Does the current applied to the surface of the head reach the ANS elements in the brain?
- Does the applied amperage stimulate the ANS and is this phenomenon accompanied by activation of endopyelotomy, serotonergic, cholinergic, and other neurotransmitter systems?
By NMR tomography, the intracranial pathways of current propagation were noninvasively investigated first. It was found that when the electrodes are sagittal (this position is conducive to the analgesia), the current is distributed in two ways: extra cerebral and intra cerebral. In both cases, the current passes through spaces containing cerebrospinal fluid. Outside the brain, the main current follows a chain of sequentially connected cerebrospinal fluid reservoirs on the basal surface of the brain.
Inside the brain, the current enters through the thin medial wall of the anterior horns of the lateral ventricles, passes through the anterior horns and enters ventricle III through the Monroe foramina. Next, the current reaches ventricle IV through the Sylvian aqueduct. Both pathways have connections through the hypothalamic structures at the bottom of ventricle III and lateral openings of Luschka in ventricle IV. With the bilateral position of the electrodes, the current does not flow to the ANS elements, and its highest density is noted in the area of the hemispheres adjacent to the area of the electrodes.
Thus, only when the electrodes are positioned so that they ensure the sagittal direction of the current, the latter has access to the main elements of the ANS (ventro medial hypothalamus, periaqueductal gray matter, and the bottom of the IV ventricle). This notion matches the results of studies where the analgesic effect occurred only with this arrangement of the electrodes.
Brainstem structures activated by the current during TES were identified in auto radiographic experiments with [14C] Deoxy Glucose (DG), which accumulates in nerve cells in accordance with their level of excitation. When pain stimulation was applied, increased accumulation of DG was observed in groups of neurons in the medulla oblongata, thalamus, and cortex, i.e. along the path of pain impulses. If a pain stimulus was during TES, then increased accumulation of DG was observed in periaqueductal gray matter and some hypothalamic structures related to the ANS. At the same time, there was no excessive accumulation of DG in the structures participating in the conduction of pain impulses. This result allowed us to conclude that under the influence of TES, due to the ANS activation, ascending pain impulses are blocked, and their entry into the cerebral cortex is prevented.
It was found that an opioid neurochemical mechanism is involved in the analgesic effect of TES. This notion was evidenced by a number of results obtained by the authors in experiments and clinical studies. First, it was noted that during and after TES, the release of an opioid peptide-β-endorphin-in the brain increased and the concentration of this peptide in cerebrospinal fluid and blood increased. At the same time, the strongest release of β-endorphin occurred when the stimulation had a frequency optimal for the analgesic effect. Second, experiments have revealed that all TES effects, including conjugated ones, were prevented or eliminated after administration of naloxone, a m-opioid receptor blocker. Third, the analgesic effect of TES did not occur in animals and people with high tolerance to morphine. Finally, the Transcranial Electroanalgesia (TEA) was significantly enhanced by substances that inhibited the activity of enzymes that break down endorphins. All these findings meet generally accepted evidence criteria for the participation of opioid peptides in the effects of ANS stimulation.
Serotonergic mechanisms of the ANS also actively take part in the implementation of the analgesic effect of TES and are probably associated with opioid drugs. Indeed, blockers of serotonin receptors or mechanisms of the release of this neurotransmitter abrogated the analgesic effect of TES, and serotonin agonists with different mechanisms of action enhanced it. This finding is important from practical point of view because serotonin agonists, unlike opioid receptor agonists, do not have narcotic properties and in the future can be used in the clinic for potentiating the analgesia caused by TES.
Recently, another mechanism of analgesic action of TPP was identified, which also has an opioid nature. Nevertheless, this mechanism is not central but rather peripheral. It turned out that opioid peptides that are strongly released into the blood during TES have a suppressive effect on peripheral skin receptors, including pain receptors. This action of TES can be especially effective against the pain associated with inflammatory processes because it is reported that the stimulated nerve endings a tan inflammation site become more sensitive to the suppressive action of endorphins.
It has been documented that the opioid connection of the ANS (Lebedev., 1987) [12] is involved in the neurochemical mechanism of TEA. With the help of metabolic mapping, it was discovered that neurons of periaqueductal gray matter and suture nuclei are activated during TEA (Lebedev., 1987) [12], and their direct electro stimulation causes analgesia, with the participation of both endogenous opioids and GABA. Many neurons that respond to GABA summation were found in ANS structures (Behbehani, 1990) [13], and the presence of cholinergic structures in the ANS was shown too. In this regard, it is possible that GABA and cholinergic projections take part in the realization of TEA.
In experiments on rats, TES and evaluation of its impact were performed by the method described earlier (Lebedev, Savchenko et al., 1988) [14].
To study the GABAergic connection, the influence of GABA antagonist picrotoxin (1.75 mg/kg) and GABA agonist baclofen (7.5 mg/kg) on the TEA severity was evaluated. After pre administration of picrotoxin, there was a slight decrease in TEA. Baclofen potentiated TEA. Along with these effects, there was a 2.5-fold increase in the pain threshold after baclofen injection and a 1.7-fold increase after picrotoxin injection. After the termination of TES in animals, there was a tendency for hyperalgesia.
To study cholinergic mechanisms, the following substances were used: M-cholinolytic scopolamine (5 mg/kg), M-cholinolytic oxotremorine (0.1 mg/kg), N-cholinolytic pirilene (0.25 mg/kg), and N-cholinomimetic nicotine (10 mg/kg). In experiments with cholinomimetics, rats of two age groups were used (I: up to 3 months, II: over 3 months) because it has been proven that the impact of these substances on the elements of the ANS manifests itself differently depending on age. Peripheral effects were blocked by M-and N-cholinolytics that did not penetrate the blood–brain barrier: metacin (2 mg/kg) and benzohexonium (5 mg/kg). In special control experiments, it was found that these substances did not affect TEA severity. In the group of cholinolytics, the most pronounced effect was exerted by scopolamine, which abrogated TEA, whereas pirilene only attenuated TEA. The influence of cholinomimetics was different. Strengthening of TEA under the influence of oxotremorine was observed only in rats of group II. On the other hand, after the administration of nicotine, hyperalgesia was observed in group I rats during TES, and in group II rats, TEA proved to be weakened. The doses of nicotine used were high enough to cause brief seizures.
Thus, GABA and cholinergic mechanisms may play a certain but probably not the main part in the implementation of TEA.
Stress: The next most important factor compromising the normal functioning of the human body is stress. A living organism is constantly exposed to the external environment, whose negative influence is increasing under the conditions of modern technological progress and civilization. Thousands of different signals enter the brain through appropriate sensory pathways, thus supporting our physiological processes or disrupting them if the defense mechanisms are not able to maintain a dynamic balance. In this context, it is customary to talk about stress as a response of the human body to a variety of external stimuli that pose a threat to homeostasis.
The term “stress” was first introduced by W. Kennon [15] to characterize stimuli that “overload” the body and disrupt homeostasis. Nonetheless, a decisive role in the confirmation of this concept was played by G. Selye [16], who first developed the theory of “general adaptation syndrome,” which has occupied the minds of researchers for more than half a century [17]. Because stress is the root cause of a huge number of diseases, this concept is often interpreted in medicine from a pathophysiological standpoint, but this approach is hardly justified. An inadequate stress response occurs only when the regulatory systems are imperfect or when the load imposed exceeds their capabilities owing to considerable magnitude or duration of stimulation. In this case, the problem of stress becomes pragmatic because it is known that by purposefully influencing the main mechanisms that regulate the stress response, researchers can achieve a sufficiently reliable adaptation and help a person to live freely under modern conditions.
One of the ways to influence altered stress systems is TES of the brain. Because the stress response is mainly controlled by the neuroendocrine system, the authors offer their vision of the approach to (and treatment of) stress.
Over the past decades, a large number of studies have been performed on brain processes caused by stress. Since the publication of the studies by Morgan et al. [18] and Hunt and coworkers [19,20], the induction of the C-FOS protein has become a popular marker for mapping of CNS neurons that are excited by various types of peripheral (mainly pain) stimulation or during convulsions [18,21–27].
Recently, it was reported that C-FOS expression or C-FOS mRNA up regulation are sensitive markers for mapping the activation of central neurons even under the action of mild stressors, such as removal of rats from a cage [28–31], holding of animals by an experimenter [32], placing them in unfamiliar conditions [18,33], forced running [34], placement of electrode clips on their ears [35], or intra peritoneal administration of inert solutions [36]. Mapping of the C-FOS protein has also been used to determine the neural circuits of the brain involved in arousal under various types of more intense stress, especially those caused by immobilization [20,37,38]. After immobilization, the largest numbers of C-FOS–positive neurons and the largest amounts of C-FOS mRNA are found in the paraventricular (RA) and Supraoptic (SO) hypothalamic nuclei, the medial part of the thalamus, and the central amygdalar nucleus [39–43]. Mapping the stress-related neurons of the brain using the C-FOS protein also gives a unique opportunity to determine the mechanism and delineate the areas affected by anti stress pharmacological and non-pharmacological effects [27,44].
In our previous studies [45], it was concluded that the therapeutic effect of a thermal power unit is related to the action of rectangular pulses having a special property. Increased levels of β-endorphin in the brain, cerebrospinal fluid, and blood; naloxone sensitivity; cross-tolerance with morphine; and potentiation with enkephalinase inhibitors have been used to show that the effect of TES in this mode is opioid in nature [14,46].
In the latter article, we investigated the effect of TES on the expression of C-FOS in certain brain structures under immobilization stress: a model that has been well studied by C-FOS mapping and enables researchers to determine weakening or strengthening of the action of stress factors.
To study the impact of TES on the neurotransmission in the forebrain, the authors have employed C-FOS expression in neurons of the forebrain under stress of different intensities. The data obtained in that paper are in good agreement with those reported earlier [20,37,38]. In previous studies, it was clearly demonstrated that stress causes an increase in the number of C-FOS–positive neurons in the cortex, medial parts of the thalamus, in the RA and SO nuclei of the hypothalamus, and in the central amygdalar nucleus. In the last three structures, C-FOS expression is regarded as a marker of the presence and intensity of stress [39–41,47–50].
The presence of C-FOS expression in the cortex under stress in our models confirms the previously published data [51,52], and expands them because of more detailed investigation of individual regions of the ISO and allocortex. It should be noted that the number of labeled C-FOS neurons even in rats with WIS was significantly lower than the number of Nissl-stained neurons. As shown quantitatively by Schleicher et al. [53] using a special index (grey scale index), the largest number of isocortex neurons is concentrated in layers I–IV. According to the grey scale index, the ratio of the number of neurons in layers I–IV, V, and VI, for example, is 1.5:1.2:1.0 in the Fr1 region, 1.3:1.2:1.0 in the Fr2 region, and 2.8:1.2:1.0 in the Pari region. According to our observations in the control subgroup of animals with MSV, in approximately the same layers of the isocortex, the ratio of labeled neurons was different and was 1.0:2.0:4.25 in the regions of Frl,1.0:3.6:4.0 in Fr2,1.0:1.59:1.5 in Pari, and 1.0:2.54:3.11 in Cg3. By contrast, in the RSA and RSG regions, this ratio of labeled neurons in distinct thirds of the cortex thickness was different because this subdivision does not correspond to the subdivision of these areas of the cortex into layers. According to Vogt et al. [54,55], layers II–IV in RSA are relatively thinner than in other areas of the cortex. In RSG, layer VI is very thin; therefore, in the deep part of the cortex in this region, the number of labeled neurons is relatively small. In general, in animals under stress, the highest expression of C-FOS was observed in the deep cortex, where most of afferent neurons are located.
As a rule, TES caused inhibition of C-FOS expression in different regions of the cortex and was more pronounced in the deep parts corresponding to layer VI. Nonetheless, the suppressive effect of the TPP was not uniform. Certain differences in the action of the TPP could be observed even in the adjacent areas of the cortex, for example, Frl and Fr2, RSA and RSG, and HL and Pari.
It is especially important to emphasize that after TES, there was no increase in the number of C-FOS–labeled neurons in any of the cortical regions. This did not happen even in the rostral cortex (Cg, Fr), where current density near the front electrode could be higher. These data are consistent with auto radiographic analysis, where it was found that TES in the same mode did not increase the basal level of absorption of 2-Deoxy-[3-N]-Glucose (DG) in the cortex of rats and prevented an increase in the absrption of DG by somatosensory regions of the cortex during nociceptive stimuli. According to Sharp et al. [27], there is a good match between mapping analyses of excited brain regions by means of DG and C-FOS.
The absence of an increase in the number of C-FOS–labeled neurons after TES in the Pir, which is directly related to olfactory bulbs, the most rostral part of the brain, also indicates that this effect does not have a direct excitatory influence on the anterior parts of the brain. Noninvasive measurements of current density and pathways inside the skull during TES by NMR imaging [56], at the same position of electrodes has revealed that the current does not propagate around the convexital surface of the cortex and probably cannot directly stimulate it. At the same time, even natural stimulation with odors can cause an increase in C-FOS expression in the brain structures associated with olfaction [30]. The observed decrease in C-FOS expression also means that TES does not have a nonspecific effect on cortical neurons, as evidenced by spreading depression, where a high level of C-FOS is detected.
Some research [57,58] indicates that TES in this mode causes a significant increase in the amount of β-endorphin in some areas of the brain, cerebrospinal fluid, and blood. In rabbits and cats, there is a decrease in the amount of β-endorphin in the pituitary gland. Analgesia caused by TES had an opioid nature because it was abrogated by naloxone, featured cross-tolerance with morphine, and was potentiated by D-amino acids that inhibit enkephalinase. In agreement with these data, it seems reasonable to assume that TES can cause an increase in the expression of C-FOS, i.e., excite Arc neurons that produce β-endorphin [59]. The same can be said about the neurons of the parvocellular part of RA containing CRF, which, by stimulating the processing of ROMs in the pituitary gland promotes the flow of β-endorphin into the blood [60–62]. Vasopressin produced by the magnocellular part of RA and SO nuclei may also play a role in the release of β-endorphin by the pituitary gland. Nevertheless, in the neurons of all these nuclei, C-FOS expression was suppressed by TES.
Thus, there is a certain contradiction between the alleged findings and the assumption following from previously obtained data. In turn, this contradiction is based on the assumption that the expression of C-FOS means the activation of neurons.
One possible explanation for this contradiction may be as follows. An increased β-endorphin release may be associated with some degree of activation of PeF neurons, some of which increase their C-FOS expression. Indeed, this nucleus contains vasopressin neurons [63], whose axons are traced to the pituitary gland [64]. In addition, the level of enkephalin immunore activity is significantly higher in this nucleus than in other hypothalamic nucleus [65].
Another possible explanation may be that the absence of C-FOS signals in the expected zones may be due to the chosen stimulation paradigm. In this work, we used the same TES paradigm (continuous stimulation at a frequency of 70 Hz for 1 h), which causes analgesia of an opioid nature in rats [66,67]. At the same time, some data suggest that rare pachytic stimulation is more effective at inducing C-FOS expression [68]. On the contrary, with prolonged stimulation or repeated stress, C-FOS expression may decrease due to habituation or desensitization [69,70]. It follows that prolonged TES, effectively used for analgesia, may not be optimal for the induction of C-FOS expression.
There are convincing reports that neurosecretory brain cells activated by long-term natural or pharmacological stimuli increase their neurosecretory activity without C-FOS expression. For example, a recently found trend isupregulation of CRF mRNA in the mediobasal hypothalamus after TES of the same frequency and duration (Kunos G, 1994, personal message) during a decrease in C-FOS expression in RA nuclei. Of note, rapid activation of the CRF gene transcription caused by a decrease in corticosterone in blood plasma takes place competitively with changes in levels of C-FOS mRNA or even before them, which raises questions about the possibility of early poststimulus influence of C-FOS on the CRF gene [71].
To resolve this contradiction, further research is required regarding the effect of TES on the expression of various “immediate-early genes” because neurons can express different groups of such genes depending on the conditions of stimulation [72,73]. Therefore, it is possible that the absence of C-FOS expression in this case does not necessarily mean that activation of hypothalamic neurons is absent [44,71].
It is also impossible to rule out the possibility that the external current applied to the areas of its highest concentration cannot have a direct suppressive effect on the discharges of excitable membranes, on the system of secondary messengers, or on other factors that usually stimulate the expression of C-FOS. The probable impact of an electric current on axoplasmic currents without effects on soma discharges of the neuron should also be taken into account in further studies [74].
Summing up the results of the experiments, it should be pointed out that TES in a mode that was previously found to be anti stress and analgesic reduces stress-induced expression of C-FOS in the neuronal nuclei of most of the studied cortical regions, medial thalamic, and some hypothalamic structures. The suppressive effect of TPP on C-FOS expression has been observed in areas with high current density and without it. We believe that the data obtained directly indicate the presence of an anti stress effect of TPP.
Regeneration of damaged neurons: Regeneration of peripheral nervous tissue is a complex process that involves various functional systems (endocrine, immune, antinociceptive, circulatory, and others). In some works, investigators have shown the possibility of accelerating the effect of the opioidergic system on the regeneration of nerve fibers using the effect of exogenous administration of a synthetic analog of leu-enkephalin, dalargin, as an example [75]. It can be assumed that endogenous opioid peptides released into the blood can also help accelerate regeneration processes.
Data on TES have been obtained that prove the opioid nature of analgesia as well as an increase in the concentration of p-endorphin in the cerebrospinal fluid and blood during electro stimulation in the analgesic mode [46,76]. Accordingly, it is possible to research the influence of TES on the regeneration of damaged nerve fibers. The authors conducted experiments on rats. After a surgical procedure, one group of animals (20 rats) was subjected to electrical stimulation using electrodes inserted subcutaneously into the forehead and behind the ears. We utilized a previously developed [67] stimulation mode in the form of a combination of a constant current of 0.8 TA and a pulse current of 0.4 GPa with a frequency of 70 Hz and a duration of 3.0 ms to 3.5 ms. These parameters of stimulation ensured the greatest analgesic effect. There were 4 sessions of exposure lasting 1 h with an interval of 3 days for 2 weeks after the surgical procedure. The second group of 20 operated animals served as a control.
Reinnervation of a rat’s foot with efferent nerve fibers was evaluated by a vestibular motor test: reflex dilution in fingers of the hind limb when the animal was abruptly lowered. This test is widely used in experiments on rats in the research on the regeneration of efferent fibers of the damaged sciatic nerve [77]. Regeneration of afferent nerve fibers has been studied by the method of withdrawing impulse activity from sciatic nerve micro beads in case of mechanical irritation of foot skin [75].
In addition, a delayed effect of TES on the functional restoration of a severed and sutured sciatic nerve was assessed 9 months after surgery. The functional state of the regenerating nerve was evaluated by registering the Total Action Potential (TAP) with subsequent calculation of nerve conduction rates as well as by registering impulse activity of fibers of the sciatic nerve during mechanical irritation of the foot skin. The results were compared with data obtained from 10 intact animals.
Evaluation of the vestibular motor reflex in the operated animals showed that the first signs of recovery of motor functions are observed after 5.0 days ± 1.0 days in the experimental group and after 9.0 days ± 1.5 days in the control group. When the impulse activity of sciatic nerve fibers was registered, it was revealed that in animals that were subjected to electrical stimulation, the first impulse responses to the strong mechanical irritation of the foot skin (tingling, prickling) take place at 14.0 days ± 1.0 days after surgery, and in control animals at 19.0; 11.0; 9.0 days after surgery. Thus, TEA accelerated the reinnervation of the foot skin by motor and sensitive nerve fibers, i.e. it stimulated the growth of regenerating nerve fibers.
Experiments conducted 9 months after the operation revealed that the conduction of the sciatic nerve was fully restored in both groups of animals. Nevertheless, the speed of conduction along the most rapidly conducting nerve fibers, as determined by the TAP registration method, was significantly lower than normal (44.5 m/s ± 0.4 m/s) and was 34.7 m/s ± 1.0 m/s and 35.0 m/s ± 1.1 m/s in the control and experiment, respectively.
Analysis of the action potentials (of stand-alone afferent nerve fibers) registered during the mechanical irritation of the foot skin indicated that the duration of the action potential in all three groups of animals ranged from 0.5 ms to 1.2 ms. As presented in the above histograms, in the intact animals, nerve fibers with an action potential duration of 0.5 ms to 0.6 ms are prevalent (0.60 ms ± 0.05 ms, mean ± SD). In control animals, 9 months after the surgery, nerve fibers with an action potential duration of 0.8 ms to 1.0 ms are prevalent (0.9 ms ± 0.05 ms). In the animals that underwent electro stimulation, there was a significant (P < 0.05) decrease in the duration of the action potential in most nerve fibers, approximating the average normal value (0.70 ms ± 0.05 ms).
The results of this study suggest that TES effects have a positive influence on regeneration processes in the peripheral nervous system. In the early stages of regeneration, the electroanalgesia apparently stimulates the growth of regenerating nerve fibers, which is manifested in accelerated initiation (by 30% on average) of reinnervation of the foot by efferent nerve fibers and by afferent ones (25% acceleration). A comparison of the parameters of reinnervation of the foot by sensory and motor nerve fibers revealed slower regeneration of sensory nerve fibers, consistently with the literature data.
The late stages of regeneration, where it is possible to assess a full recovery of the damaged nerve functions, are especially relevant to the clinic. Experiments were conducted 9 months after the operation, because by this time, regenerating nerve fibers reach the target tissues, and their myelination is almost completed [31]. In the analysis of the TAP of the sciatic nerve soma, it was found that the speed of conduction did not differ significantly between the experiment and control. Under our conditions of withdrawal, we can only assess the speed of conduction by the most rapidly conducting (most likely motor) nerve fibers. Probably, at this stage of regeneration, the electrical effect does not influence the speed of conduction through the efferent nerve fibers.
It is known [78] that injuries more severely affect sensory nerve fibers, because of their greater dependence on retrograde transport of neurotrophic factors from a target tissue. Perhaps this is why regeneration processes in sensory fibers are slower. The final stage of regeneration of the damaged nerve is restoration of the sensitivity of the denervated tissue. Accordingly, the analysis of action potentials registered in single sensory fibers was carried out in response to mechanical irritation of the foot skin. As the main parameter, action potential duration was chosen, which depends on functional properties of afferents and above all on the speed of conduction along the nerve fiber. A comparison of action potential duration between intact animals and two groups (control and experiment) of operated animals showed that TEA contributes to better restoration of functional properties of the damaged afferents.
Consequently, TEA has a positive impact on functional restoration of a damaged rat sciatic nerve by stimulating the growth of regenerating nerve fibers and their functional maturation. The totality of evidence supporting the opioid nature of electroanalgesia [14,46,76], suggests that its effect on regeneration is related to an increase in the systemic level of endogenous opioid peptides. Opioid peptides and their receptors are widely distributed both in the CNS and at the periphery [79–82]. Therefore, it is not surprising that they are multifunctional, as noted by many authors [79,83]. In addition to the well-known analgesic effect, it has been established that opioid peptides take part in the regulation of immunity, have a universal anti stress effect, participate in the regulation of cardiovascular and respiratory systems, and affect repair processes [82,84–89]. Data from experiments on tissue culture [90,91] indicate that opioid peptides have a neurite-stimulating and trophic effect as well as accelerate the proliferation and migration of glia. It can be assumed that under the conditions of a complete organism, the positive effect of opioids on the regeneration of peripheral nerves consists of several components: an effect on the resistance of neurons to injury, stimulation of neurite growth, and acceleration of proliferation and outgrowths of glial cells in the growth zone. In addition, the normalizing effect of opioids on microcirculation and lymph flow—and their immunomodulatory and anti stress properties—can play a substantial part in the regeneration of nervous tissue.
The antitumor effect: The system of endogenous opiates (i.e., opiate receptors and their ligands) plays a considerable role in the regulation of tumor growth [92]. The presence of opiate receptors has been demonstrated on the cell membrane of small-cell lung cancer cells in humans [93], on human SK-N-MC neuroblastoma cells [94], mouse neuroblastoma S20Y cells [95], and on many other benign and malignant tumor cells of mesodermal, ectodermal, and endodermal origin in humans and animals.
A study on the properties of opiate receptors has uncovered specific, high-affinity, saturated binding of various Opioid Peptides (OPs) on the cell membranes of these tumors. Radio immunological methods revealed the presence of β-endorphin and met-enkephalin in these tumors. Met- and leu-enkephalin were detected in tumor tissue immunocytochemically. They are associated with the perimembrane cytoplasm of tumor cells. All these data allow us to conclude that the presence of OPs and their receptors is a fundamental characteristic feature of human and animal tumors [96].
Binding of opiate receptors to their corresponding ligands introduced externally, in most cases, leads to inhibition of tumor growth in tissue culture. For example, met-enkephalin suppresses the proliferation of mouse neuroblastoma S20Y cells [97], mouse neuroblastoma SK-N-MC and human neuroblastoma cells [98], and human fibrosarcoma NT-1080 cells [99]. On the other hand, there are reports on the stimulation of the growth of human V-373-MG astrocytoma cells by met-enkephalin [98].
Of great interest are data on the regulation of tumor growth in tissue culture by naltrexone, a potent opiate receptor blocker. Naltrexone stimulated the growth of human astrocytoma 373-MG cells, mouse S20Y neuroblastoma cells [97], mouse n-114 neuroblastoma cells, NT-1080 fibro sarcoma cells, and human SK-N-MC neuroblastoma cells [100]. By contrast, naltrexone suppresses the growth of human SK-N-MC neuroblastoma cells [98].
The apparent contradiction between the last two results obtained by different authors on the same tumor type is easily explained because it is known that the antitumor effect of OPs and their antagonists depends on their concentration and the number of tumor cells. For example, the growth of human K-N-MC neuroblastoma cells transplanted into athymic nude mice in the amount of 2.5 × 106 cells was suppressed by daily injections of naltrexone at a dose of 0.1 mg/kg but was enhanced by daily injections of this substance at 10 mg/kg [95]. Naltrexone at a dose of 0.1 mg/kg also suppressed the growth of S20Y neuroblastoma in mice [95] and human NT-29 rectal cancer cells [101] transplanted into athymic nude mice in the amount of 106 to 2.5 × 104 tumor cells. It should be noted that the use of naltrexone at 0.1 mg/kg led to the blockade of opiate receptors for 6 hours to 8 hours per day, but in the remaining 18 hours to 6 hours, the sensitivity of the receptors to OPs, in particular to met-enkephalin, increased 2-fold [102].
Met-enkephalin at 0.5 mg/kg to 30 mg/kg slowed the growth of S20Y neuroblastoma that was transplanted into normal A/Jax mice in an amount of 103 to 2.5 × 104 cells, and this effect was blocked by naloxone [92]. Met-enkephalin at 3 mg/kg inhibited the growth of melanoma B-16 cells transplanted into mice in the amount of 106 cells (the effect was also blocked by naloxone [104]). In general, met-enkephalin is currently considered an agent that inhibits the growth of tumor cells [100] owing to an interaction with opiate receptors located on their membrane.
The system of endogenous opiates performs a major function in the regulation of the body’s immune responses [84]. There is evidence that opiate receptors are present in immunocompetent cells [104], including natural killer cells [105], which implement antitumor immunity. Binding of opiate receptors to their endogenous ligands, in particular β-endorphin increases the activity of natural killers [105,106], and this phenomenon is blocked by naloxone.
Thus, to activate the endogenous opiate system and its effect on antitumor activity, TES can be employed in a mode that leads to a several fold increase in the concentration of β-endorphin in the blood [107]. The authors of that study investigated the effects of a synthetic analog of OP leu-enkephalin (dalargin) and TES on the growth of transplanted tumors in mice and rats when applied separately or in combination with chemotherapeutic drugs cyclophosphane and sarcolysin.
Walker 256 carcinosarcoma served as the tumor model. When TES was used in the mode (70 Hz and 3 ms) leading to the maximum allocation of OPs, there was pronounced inhibition of tumor growth (by 33% to 43%) at different time points of observation. In the next series of assays, it was found how deviation of any one of the TPP parameters from the optimal mode can affect its antitumor effect.
It was demonstrated that when the pulse duration was fixed at 3 ms, the deviation from the optimal frequency to 90 uti/s leads to a significant increase in the growth of Walker’s carcinosarcoma (up to 61%) on the 9th day and afterwards. By day 13, the mass of the tumor in the control groups and when the frequencies of 50 uti/s and 90 uti/s were employed was approximately the same, i.e. there was no noticeable stimulatory effect. TES at an optimal frequency of 70 Hz, but with reduced pulse duration of up to 1 ms, stimulated the tumor growth by 48% to 60% on different days of the experiment. If the TES was carried out at the optimal frequency of 70 Hz with longer pulses (5 ms), then there was 12% to 40% inhibition of tumor growth, which was significant under one of the conditions after transplantation. In this case, the antitumor effect was close to the effect of TES at 70 Hz and 3 ms but did not exceed it, because in the latter case, the tumor growth inhibition was detectable during all periods of observation. Thus, the authors showed that the maximum inhibitory effect on the growth of Walker’s transplanted carcinosarcoma manifests itself in the optimal mode of TES, which is necessary for stimulating opioid mechanisms.
In the next series of experiments, the effects of the thermal power unit were studied in the optimal mode, but before not after the Walker’s carcinosarcoma was transplanted. With this scheme of the experiment, there was pronounced significant inhibition of tumor growth by 33% to 45%, which was not different from the effect of TPP after the transplantation. The obtained results indicate that the action of TES is apparently directed at the antitumor defense system of the body.
To characterize the spectrum of the antitumor effects of TES, the authors studied its influence on the growth of three more transplanted tumor types in rats. It was found that TES, when used in the optimal mode on the next day after the transplantation, significantly inhibits the growth of Pliss lymphosarcoma (by 42% to 46%) and sarcoma (by 45% to 60%) and significantly inhibits the development of an ascitic ovarian tumor in rats transplanted with 106 tumor cells. At the same time, the effectiveness of the antitumor action of TES on the affected ovarian tissue manifested itself both in a significant decrease in the volume of ascites (by 43.6%) and in a significant decrease in the volume of a dense sediment of tumor cells (by 35%).
There was a tendency for a delay in tumor formation in the rats exposed to TES after transplantation of Pliss lymphosarcoma (by 3%), sarcoma 45 (by 19%), and Walker’s carcinosarcoma (by 4%) (The effect was prevented by naloxone). Of note, there was a significant increase in the life expectancy of the first deceased rat in the groups subjected to TES after transplantation of ascitic ovarian tumors (by 111%), Pliss lymphosarcoma (by 42%), sarcoma 45 (by 54%), and Walker’s carcinosarcoma (by 46%) (naloxone reduced this effect to 38%) in comparison with the control.
Summing up all the above, we can conclude that TES in the optimal mode leads to reliable inhibition of the growth of transplanted tumors of various types as well as to an increase in the life expectancy of tumor-bearing animals. At the same time, this TES mode matches in all parameters the mode that leads to the maximal activation of opioid mechanisms, in particular, to an increase in the concentration of β-endorphin and met-enkephalin in the rat cerebrospinal fluid and to a maximum increase in the concentration of β-endorphin in the blood of rats. At the same time, deviation from this optimal mode in at least one parameter leads either to the disappearance of the antitumor effect or even to the stimulation of tumor growth. Direct evidence of the participation of the endogenous opiate system in the antitumor effect of TES is the results of experiments with naloxone.
In accordance with the above data, the TPP with an optimal frequency of 70 imp/s and pulse duration of 3 ms significantly inhibits the growth of Walker’s carcinosarcoma, by 30% to 36%. Naloxone in a total dose of 12 mg/kg, administered during the TPP, significantly reduced this antitumor effect. This finding indicates that there is an opioid component in the mechanism of the antitumor effect of TES, and it seems that β-endorphin and opioid receptors are involved. These data are in good agreement with the results of several studies on the strengthening of the immune defense system by opioid receptors, namely, increased mobility of lymphocytes [108], enhancement of cytotoxic properties of natural killer cells [105,106,109], production of kinins [106], antibody synthesis, and the reversal of these effects by naloxone [105,106,108,109].
In addition, the direct action of an OP released during TES, in particular met-enkephalin, on the corresponding receptors of tumor cells cannot be ruled out. Meanwhile, it is known that the acceleration or deceleration of tumor growth during its autocrine regulation depends on the concentration of OP-expressing cells [110]. It is possible that with TES, their concentration reaches the level necessary to inhibit tumor growth. The fact that naloxone does not completely abrogate the antitumor effect of TES can be explained as follows. It is known that there are receptors for β-endorphin that are not blocked by naloxone on lymphocytes, including spleen lymphocytes of rats involved in antitumor defense [111,112], that and the immunostimulatory effect of β-endorphin in vitro is reduced but not completely eliminated by naloxone [113].
Electrical stimulation in parkinson’s disease: A number of studies suggest that chronic electrical stimulation of deep brain structures belonging to basal ganglia, such as motor centers of the thalamus, globus pallidus (pale ball), and later the Subthalamic Nucleus (STN), effectively improves motor performance in Parkinson’s Disease (PD) [114–119]. Thalamic stimulation in the ventral intermediate nucleus can significantly alleviate contra lateral tremor, while the pale ball and the STN are effective targets for reducing rigidity and akinesia, while stimulation of the STN has more consistent antikinetic effects, a greater reduction in the interaction of anti parkinsonian drugs, and lower required stimulus intensity [120]. In particular, high-frequency (> 100 Hz) stimulation of the STN influences a number of functional outcomes related to lower extremities (walking), upper extremities (proximal and distal movements, including hand writing), and muscles of the tongue (speech). Improvements in monotonous mechanical labor such as treadmill walking and ground walking have been shown (increasing stride length and/or increasing frequency [121,122], torso kinematics, lower-limb joint kinematics, and joint dynamics (normalization of moments and force) [123]. The identified benefits for upper-limb function affect slow downward movements, ballistic forearm movements, and isometric contractions of the forearm muscles (increased speed and strength); however, they are fairly simple for tasks involving more subtle wrist/hand movements (moderate increases in the rate of grasping, tipping, and the peg board test as well as evenness and accuracy of hand writing [124–126]. It has been demonstrated that changes in speech structure are even less obvious (a slight increase in the level of sound pressure and variability of the main frequency [127,128] or negative (deterioration of verbal fluency [129]).
Thus, an intervention in the activity of the STN in patients with PD through high-frequency electrical stimulation affects a range of motor tasks but tends to influence mainly axial, proximal, and locomotor movements, that is, those that are controlled by relatively lower hierarchical levels of the motor system.
Bilateral electrical stimulation is also used for the treatment of PD. Bilateral deep brain stimulation of the STN has proven to be an effective treatment for patients with PD both short- and long-term, thereby leading to good control of all major symptoms (rigidity, bradykinesia, and tremor) and of clinical motor complications (motor fluctuations and dyskinesia) [128,130–134]. In addition, studies involving instrumental motion analysis have revealed significant improvements provided by bilateral deep brain stimulation of the STN for advanced postural adjustments prior to the initiation of walking [135], kinematics and kinetics of joints of the lower extremities during steady walking [123], and locomotor synergies of upper extremities [136], which are usually affected at the stage of PD progression.
Although bilateral deep brain stimulation of the STN is considered a relatively safe procedure, in recent years, there was growing interest in unilateral and phased deep brain stimulation of the STN (i.e., implantation of two electrodes during two surgical procedures separated in time), which poses a lower surgical risk and fewer postoperative complications such as cognitive dysfunction [137–140]. Alberts and many others [139], for example, have reported a decrease in cognitive and motor functions when examining patients subjected to two-way stimulation but not with one-way deep brain stimulation of the STN.
In addition, several studies have shown that unilateral deep brain stimulation of the STN is also effective against the symptoms of PD by improving the overall scores of the Unified PD Assessment Scale (15) by~20% to 40% (16 to 20). Improvement of motor symptoms of PD as a result of unilateral deep brain stimulation of the STN is mainly observed on the contra lateral side of the body toward the stimulated STN; however, a number of authors have also demonstrated the corresponding ipsilateral effect of unilateral deep electrical stimulation of the STN. In addition, possible dominance of right-hand motor networks for gait control was proposed in a recent study by Lizarragi et al. [141], which evaluated the effects of bilateral, right-sided, or left-sided deep brain stimulation on walking.
Judging from the above studies, ESM is an effective and fairly universal way to treat various diseases.